Tao Wentao, Du Baojie, Ma Shuai, Chen Rui. Ant Pupae from Myanmar Amber Reveal the Evolutionary Process of Cocoon-Spinning Behavior in Formicidae[J]. Journal of Gems & Gemmology, 2023, 25(4): 120-135. DOI: 10.15964/j.cnki.027jgg.2023.04.011
Citation:
Tao Wentao, Du Baojie, Ma Shuai, Chen Rui. Ant Pupae from Myanmar Amber Reveal the Evolutionary Process of Cocoon-Spinning Behavior in Formicidae[J]. Journal of Gems & Gemmology, 2023, 25(4): 120-135. DOI: 10.15964/j.cnki.027jgg.2023.04.011
Tao Wentao, Du Baojie, Ma Shuai, Chen Rui. Ant Pupae from Myanmar Amber Reveal the Evolutionary Process of Cocoon-Spinning Behavior in Formicidae[J]. Journal of Gems & Gemmology, 2023, 25(4): 120-135. DOI: 10.15964/j.cnki.027jgg.2023.04.011
Citation:
Tao Wentao, Du Baojie, Ma Shuai, Chen Rui. Ant Pupae from Myanmar Amber Reveal the Evolutionary Process of Cocoon-Spinning Behavior in Formicidae[J]. Journal of Gems & Gemmology, 2023, 25(4): 120-135. DOI: 10.15964/j.cnki.027jgg.2023.04.011
Tao Wentao: Wentao Tao (1996-), male, master of science, mainly engages in the research of insect evolution revealed by Burmese amber. E-mail: 2120190987@mail.nankai.edu.cn
Corresponding author:
Rui Chen (1987-), male, doctor, mainly engages in the research of biological evolution. E-mail: chenrui@ioz.ac.cn
Many extant species spin cocoons before their pupal stage, which can protect them from predators, pathogens, and fluctuant environmental factors in nests. However, owing to the scarcity of ant cocoon fossils, little is known about the origin and evolution of this behavior in Formicidae. Here, we describe two ant pupae specimens from Myanmar amber. The presence of cocoons in these Mesozoic fossils suggests the ancestral origin of spinning cocoon behavior; based on our phylogenetic analysis and ancestral state reconstruction, this behavior was secondarily lost at least six times in Formicidae, which could be ascribed to the transition of nesting and feeding habits, as well as the development of eusociality. Moreover, the first specimen preserved worker ants with empty cocoons simultaneously, indicating garbage removal or brood care behavior, which improves our understanding of their eusociality in the Cretaceous.
With over 14 600 described species living in nearly all terrestrial ecosystems (Bolton, 2021), ants are considered one of the most successful insect groups worldwide. These widespread hymenopteran insects usually build nests in soil or rotten wood and live in crowded societies (Pull C. et al., 2018). Compared with solitary lifestyles, social living not only brings a great deal of benefits to ants, such as efficient brood care, collective foraging, and defense, but also causes problems (Cremer et al., 2007). For instance, with a larger number of individuals living in an environment that contains a high abundance and diversity of pathogenic microorganisms, infectious diseases are more likely to spread between community members, especially among the thin-cuticled larvae and pupae (Cremer et al., 2007; Tragust et al., 2013). Thus, as in many other hymenopteran insects (e.g., wasps and bees), before the pupal stage, many extant ant’s larvae will produce a cocoon to help them stay away from predators, parasitoids, pathogens, and external fluctuating environmental factors (Armitage et al., 2012; Sarzetti et al., 2019).
In the last 10 years, after the rise of the concept of “social immunity” (Cremer et al., 2007), many experiments have been conducted to explore cocoons’ functions in ant society. Their results showed that ant pupae with cocoons have a lower probability of fungal infection, and cocoons can also protect pupae from antimicrobial acids sprayed by other worker ants during the nest sanitization process (Pull C. et al., 2018; Pull, C. D. et al., 2018; Tragust et al., 2013).Even though the cocoon plays an important role in ant social immunity, some clades of Formicidae lack this feature, which deserves further consideration. For example, all members of the subfamilies Leptanillinae, Dolichoderinae, Pseudomyrmecinae, and Myrmicinae do not spin silk cocoons before the pupal stage (Armitage et al., 2012). Many researchers attribute the absence of pupal cocoons in these clades to secondary loss and argue that a protective cocoon is an ancestral feature of Formicidae, and is even primitive in Hymenoptera (Campbell et al., 2014; Peters et al., 2017; Sarzetti et al., 2019). However, it is difficult to explain why Leptanillinae, one of the most basal lineages of the extant ant, has a naked pupal stage (Armitage et al., 2012; Peters et al., 2017; Sarzetti et al., 2019). Thus, in the latest year, some researchers started to use phylogenetic methods to reconstruct the ancestral state of cocoon in Formicidae, and their results suggested that the loss of cocoon status is associated with the presence of metapleural gland and worker polymorphism (Greer & Moreau, 2021).
The molecular divergence dating analysis of crown ants suggests that ants originated from the Late Jurassic to Early Cretaceous, but the oldest explicit ant fossil is relatively younger than that (Brady et al., 2006; Moreau et al., 2006). To date, ants have been found in 5 major amber deposits of the Cretaceous: France (100 Ma), northern Myanmar (99 Ma), New Jersey, USA (92 Ma), north Siberia (85 Ma), and west Canada (78 Ma), ranging from approximately 100 to 78 million years ago (LaPolla et al., 2013). Nearly all fossil ants from the Cretaceous belong to stem groups, which mainly consist of 2 subfamilies: Sphecomyrminae and Haidomyrmecinae (Barden, 2017; Perrichot et al., 2020). Similar to crown-group taxa, these Cretaceous stem-group ants are also eusocial, which can be reflected in their morphological features (e.g. the metapleural gland and differentiation of castes) and behaviors (e.g. aggregation of worker ants and mating flight) (Barden & Grimaldi, 2016; Krynicki, 2013). Thus, it could be assumed that stem-group ants may also face some sanitation problems due to their eusociality, which requires them to take action to curb the spread of diseases in their nests.
Here, we report 2 ant pupal specimens preserved in mid-Cretaceous Myanmar amber. The first specimen contains 3 empty pupal cocoons and 2 worker ants, which belong to the same species, and is described as Gerontoformica sp. here. In the second specimen, according to the existence of petiole structure and similarity in cocoon’s texture, we suggest that it also belongs to Formicidae, probably a member of Sphecomyrminae. However, considering that this pupa is covered by a silk cocoon, which makes it difficult to observe, we cannot provide further identification. To the best of our knowledge, this is the second-time description of the holometabolous development process of these extinct stem-group ants(Boudinot et al., 2022), and it also suggests that the most basic lineage of ants would produce cocoons during their pupal stage. Furthermore, to determine the evolutionary patterns of cocooning behavior in ants, we used 43 morphological characters mostly presented by (Barden & Grimaldi, 2016), and re-selected some species to build up a Formicidae phylogenetic tree. Compared with previous research, in order to explore the early origination of spinning cocoon behavior in ants, we paid more attention to the basic lineages and fossil ants, and our ancestral state reconstruction result shows that this behavior was lost secondarily at least six times in Formicidae at subfamily level. Finally, as to the reason why this behavior was lost secondarily, our statistical analysis indicated that there is a close relationship to 3 ecological factors: nesting habits, diet and eusocial level.
MATERIALS AND METHODS
Materials and Photography
The new amber specimens, STJ-452 and STJ-446, were collected from the Angbamo site, Tanai Township, Myitkyina District, Kachin Province, Myanmar, and specific localities were depicted in Cruickshank & Ko (2003). According to the biostratigraphic analysis, Myanmar amber has been dated to originate in the late Albian-Cenomanian age, around 105 to 95 Ma. This estimation has been refined by the radiometric dating of zircons from the volcaniclastic matrix surrounding the amber to approximately 98.8 ± 0.6 million years (Shi et al., 2012). This kind of amber is supposed to originate from the resin of a kind of conifer, probably belonging to either the family Cupressaceae or Araucariaceae, which used to live in moist tropical regions (Grimaldi et al., 2002; Westerweel et al., 2019). These two amber specimens are golden brown, translucent with some thin flows inside, and both were cut, ground, and polished. Specimen STJ-452: 26 × 17 × 5 mm (length × width × height), and 1.28g weight. Specimen STJ-446: 22 × 12 × 3 (length × width × height), and 0.58g weight. Type specimens were deposited in the Paleo-diary Museum of Insect Amber, Fujian, China. The amber specimens were examined using a LEICA M125 C dissecting microscope. Photographs were taken using a LEICA MC 190 HD fitted to the LEICA M125 C stereomicroscope. Stacks of photographs taken at different depths of the field-of-view were merged using Helicon Focus 6. Figures were prepared in Adobe Photoshop CC and Adobe Illustrator.
Materials Availability
The specimens studied are accessioned at the Paleo-diary Museum of Insect Amber, Fujian, China, with the specimen numbers STJ-452 and STJ-446. They are presently on display for public viewing.
Phylogenetic Analysis
We used 43 morphological characters mainly presented in (Barden & Grimaldi, 2016), and re-selected some species (41 in total) to cover all subfamilies of Formicidae, to set up the phylogenetic tree. Unknown characters were coded as “?”, while inapplicable characters were coded as “-”. The data matrix is presented in Table 1. The data set was edited in Mesquite v. 3.6 and analyzed using TNT v. 1.5. Searches were performed under both equal and implied weighting (K value = 6.875) as in (Goloboff, 1993). We used “traditional” and “new technology” searches (with sectorial searches, drifting, fusing, and ratchet functions) to obtain consensus trees (under strict rule). To incorporate hypotheses generated from molecular phylogenetics, the topology structure of crown-group ants was then constrained in the result of a traditional search to reflect the subfamilies’ relationships presented in Borowiec et al.(2019).This constraint was enforced only after weighted and unweighted unconstrained analyses found that Cretaceous ants were not recovered within the clade of the crown-group Formicidae.Node support values were evaluated with Bremer support and bootstrap resampling (setting for Bremer support values calculation: tree bisection reconnection from existing trees, retain trees suboptimal by 10 steps; setting for bootstrap values calculation: traditional search, number of replicates: 1 000). Character states were mapped on a strict consensus tree using WinClada v. 1.00.08.
Table
1.
Morphological character matrix used in the phylogenetic analysis.
The Morphological Characters Coded for Phylogenetic Analysis
Descriptions of characters 1-42 are quoted from (Barden & Grimaldi, 2016), and character 43 see Table 1: 1.Orientation of head: (0) hypognathous (1) prognathous; 2. Mandibular orientation: (0) parallel to axial plane of head (1) perpendicular to axial plane of head; 3. Anterior margin of clypeus with row of peg-like denticles: (0) absent (1) present; 4. Labral pegs: (0) absent (1) present; 5. Clypeus development: (0) highly reduced (1) a clearly visible sclerite (2) extending to vertex of head; 6. Clypeal suture syndrome: (0) clypeal sulci absent (1) highly reduced (2) developed; 7. Position of antennal sockets: (0) posterior to clypeus (1) anterior; 8. Clypeal process: (0) absent (1) present; 9. Torulus syndrome: (0) torular sclerite not visibly raised (1) developed as shield covering antennal base (2)developed and fused with frontal lobe; 10. Frontal lobe: (0) absent (1) single lobe not derived from frontal carina (2) present and bilobed (3) originatingas broad and bifurcating; 11. Frontal lobe syndrome: (0) bulbous (1) pinched-in posteriorly; 12. Scape length: (0) equal to all other flagellomeres individually (1) larger than any other antennal segmentsbut less than 0.25× antenna length (2) greater than 0.25× total antenna length; 13. Antennal segments: (0) less than 12 (1) equal to 12 (3) more than 12; 14. Ocelli: (0) absent (1) present; 15. Compound eye: (0) absent (1) present as single ommatidium (2) present as compound facets; 16. Development of posterior portion of pronotum: (0) narrow strip (1) developed; 17. Pronotal lobe: (0) absent (1) present; 18. Propleuron: (0) reduced, not visible in lateral view (1) developed, visible in lateral view; 19. Pro-mesonotal suture: (0) absent (1) present and flexible (articulating) (2) present and rigid (notarticulating); 20. Metanotum: (0) Distinct sclerite (1) reduced to a groove (2) obliterated; 21. Propodeal suture: (0) absent 1) present; 22. Mesopleural sutures: (0) absent/obliterated (1) anterior suture present only (2) posterior suture present only (3) both sutures present; 23. Metapleural gland: (0) absent (1) present; 24. Trochantellus: (0) absent (1) present; 25. Pretarsal claw: (0) simple (1) with subapical tooth (2) pectinate (3) with basal tooth; 26. Petiole: (0) absent (1) present; 27. Petiole peduncle: (0) absent (1) present; 28. Petiole development: (0) no dorsal expansion, cylindriform (1) gradually sloped, rounded dorsally, domeshaped (2) anteroposteriorly flattened, tall and broad (3) narrowed anteroposteriad into a scale-like structure; 29. Posterior face of petiole: (0) narrowed (1) broadly attached to gaster; 30. Petiole fusion: (0) fused tergosternally (1) no fusion, suture visible; 31. Helcium: (0) concealed (1) exposed, extends to reach petiole; 32. Third abdominal segment: (0) first gastral segment (1) postpetiole present; 33.Sternal projection on abdominal segment 3: (0) absent (1) present; 34.Constriction between abdominal segments 3 and 4 (not postpetiole): (0) absent (1) present dorsally (2)present ventrally (3) present dorsally and ventrally; 35.Abdominal segments 3 and 4: (0) tergosternally fused (1) no such fusion (2) 2+3 fused (3) only 3 fused; 36.Sting: (0) absent (1) present; 37.Acidopore: (0) absent (1) present; 38.Clypeal denticle structure: (0) largely subcuticular (1) extending well outside of cuticle with tapered apex (2) extending well outside of cuticle with flattened apex; 39.Metapleural gland opening: (0) gaping (1) opening a narrow slit; 40.Frontal carina: (0) absent (1) longitudinal sculpturing leading from the clypeus posteriad (2) circularsculpturing leading around antennal base; 41.Pygidial teeth: (0) absent (1) present; 42.Mandibular shape: (0) scythe-like with single apical tooth (1) simple with bidentate apex (2) scythe-likewith multiple teeth (3) broad, flattened apex (4) broad and triangular throughout (5) flattened basally with dramatically rounded and pointed apical tooth (6) cup-like with setose inner margin; 43.Pupal state: (0) naked; (1)polymorphic; (2) covered by cocoon.
Ancestral State Reconstruction
To evaluate the evolutionary process of cocoon-spinning behavior, we performed ancestral state reconstruction using the “Trace Character Over Trees” option in Mesquite v. 3.6 (Maddison, W. & Maddison, D., 2009). Parsimony and likelihood approaches were used to reconstruct the ancestral state based on the strict consensus tree that we obtained in the previous step. Subsequently, the average frequencies across trees were calculated, and trees were exported as PDF files, which were finally identified in Adobe Photoshop CC and Adobe Illustrator. In this procedure, because data on the character state of cocoons were rare at the species level, we used the statistical results at the subfamily level to reconstruct the ancestral state of this behavior. All known states of ant cocoons at the genus level are shown in Table 2. In addition, based on this, we also conducted a statistical analysis in Excel about the relevance between the pupal cocoon state and the nesting habits, diet, colony size, and forging behavior of ants.
Table
2.
Documentary of cocoon state on the genus level, coded with the nesting habit, diet, colony size and foraging behavior in Formicidae*
Genus
Cocoons State1
Nesting Habit2
Diet2
Mean Colony Size2
Forging Behavior2
Leptanillinae (0/0/1-8)
Leptanilla
Absent
Ground
Predator
150
Cooperative
Proceratiinae (1/1/1-3)
Discothyrea
Present
Ground
Predator
73
?
Proceratium
Polymorphic
Arb&Gro
Predator
13-200
Solitary
Probolomyrmex
Absent
Ground
Predator
20
Solitary
Amblyoponinae (4/0/0-9)
Apomyrma
Present
Ground
Predator
< 100
?
Amblyopone
Present
Ground
Predator
12
Cooperative
Mystrium
Present
Ground
Predator
< 200
Cooperative
Onychomyrmex
Present
Ground
Predator
50-400
Cooperative
Ponerinae (13/0/0-47)
Diacamma
Present
Ground
Predator
30-60
Solitary
Dinoponera
Present
Ground
Omnivore
30-60
Solitary
Harpegnathos
Present
Ground
Predator
35-225
Solitary
Anochetus
Present
Arb&Gro
Predator
100-400
Solitary
Cryptopone
Present
Ground
Predator
30-999
?
Hypoponera
Present
Ground
Predator
21-1 500
?
Leptogenys
Present
Ground
Predator
10-50 000
Cooperative
Myopias
Present
Ground
Predator
< 100
Cooperative
Odontomachus
Present
Arb&Gro
Predator
18-10 000
Cooperative
Ophthalmopone
Present
Ground
Predator
20-800
Solitary
Pachycondyla
Present
Ground
Omnivore
9-800
Cooperative
Platythyrea
Present
Arb&Gro
Predator
19-500
Solitary
Ponera
Present
Ground
Predator
30-60
Cooperative
Genus
Cocoons State1
Nesting Habit2
Diet2
Mean Colony Size2
Forging Behavior2
Paraponerinae (1/0/0-1)
Paraponera
Present
Arb&Gro
Omnivore
>2 000
Cooperative
Dorylinae (7/3/2-27)
Acanthostichus
Present
Ground
Predator
?
Solitary
Aenictus
Polymorphic
Ground
Predator
850 000
Cooperative
Cerapachys
Polymorphic
Ground
Predator
75-200
?
Dorylus
Absent
Ground
Predator
100 000
Cooperative
Eciton
Present
Ground
Predator
100 000
Cooperative
Sphinctomyrmex
Present
Ground
?
?
?
Simopone
Present
Arboreal
Predator
?
?
Syscia
Absent
Ground
?
?
?
Labidus
Present
Ground
Predator
1 000 000
Cooperative
Neivamyrmex
Polymorphic
Ground
Predator
110 000
Cooperative
Nomamyrmex
Present
Ground
Predator
1 000
Cooperative
Lioponera
Present
Arb&Gro
Predator
?
Cooperative
Aneuretinae (1/0/0-1)
Aneuretus
Present
Ground
Omnivore
65-75
Cooperative
Dolichoderinae (0/0/7-28)
Azteca
Absent
Arboreal
Omnivore
100-1 000
Cooperative
Bothriomyrmex
Absent
Ground
Omnivore
?
?
Dolichoderus
Absent
Arb&Gro
Omnivore
25-1 000
Cooperative
Iridomyrmex
Absent
Ground
Omnivore
79 000
Cooperative
Leptomyrmex
Absent
Ground
Omnivore
350
?
Linepithema
Absent
Arb&Gro
Omnivore
>1 000
Cooperative
Tapinoma
Absent
Arb&Gro
Omnivore
100-5 000
Cooperative
Myrmeciinae (2/0/0-2)
Myrmecia
Present
Ground
Omnivore
15-1 500
Cooperative
Nothomyrmecia
Present
Ground
Omnivore
50-70
Solitary
Pseudomyrmecinae(0/0/3-3)
Myrcidris
Absent
Arboreal
Herbivore
100
?
Pseudomyrmex
Absent
Arb&Gro
Omnivore
200-10 000
Solitary
Tetraponera
Absent
Arboreal
Omnivore
7 000
Cooperative
Formicinae (6/4/5-51)
Acropyga
Polymorphic
Ground
Herbivore
?
?
Brachymyrmex
Present
Arb&Gro
Omnivore
22-125
Cooperative
Camponotus
Present
Arb&Gro
Omnivore
55 000
Cooperative
Colobopsis
Absent
Arboreal
Herbivore
?
?
Formica
Polymorphic
Ground
Omnivore
150 000 000
Cooperative
Gesomyrmex
Absent
Arboreal
Omnivore
150
?
Gigantiops
Present
Ground
Omnivore
>1 000
Solitary
Lasius
Polymorphic
Ground
Omnivore
1 250 000
Cooperative
Myrmoteras
Present
Ground
Predator
8-22
Solitary
Myrmecocystus
Present
Ground
Omnivore
15 000
?
Oecophylla
Absent
Arboreal
Omnivore
500 000
Cooperative
Paratrechina
Absent
Arb&Gro
Omnivore
2 000
Cooperative
Plagiolepis
Present
Arb&Gro
Omnivore
< 5 000
?
Polyrhachis
Polymorphic
Arb&Gro
Omnivore
60-90 000
?
Prenolepis
Absent
Ground
Omnivore
3 370
Cooperative
Myrmicinae (0/0/28-148)
Acromyrmex
Absent
Arb&Gro
Herbivore
1 000-10 000
Cooperative
Aphaenogaster
Absent
Ground
Omnivore
100-300
Cooperative
Apterostigma
Absent
Ground
Herbivore
30-150
Cooperative
Atta
Absent
Ground
Herbivore
1 750 000
Cooperative
Cardiocondyla
Absent
Ground
Omnivore
>50
Cooperative
Crematogaster
Absent
Arb&Gro
Omnivore
40-29 000
Cooperative
Cyphomyrmex
Absent
Ground
Herbivore
< 500
Solitary
Eutetramorium
Absent
Ground
Predator
1-50
Cooperative
Formicoxenus
Absent
Ground
Omnivore
< 100
?
Harpagoxenus
Absent
Ground
Omnivore
10
Cooperative
Leptothorax
Absent
Ground
Omnivore
20-60
Cooperative
Genus
Cocoons State1
Nesting Habit2
Diet2
Mean Colony Size2
Forging Behavior2
Messor
Absent
Ground
Herbivore
100-1 000
Cooperative
Monomorium
Absent
Ground
Omnivore
>10 000
Cooperative
Mycocepurus
Absent
Ground
Herbivore
< 1 000
?
Myrmecina
Absent
Ground
Omnivore
24-100
?
Myrmica
Absent
Ground
Predator
86-3 000
?
Myrmicocrypta
Absent
Ground
Herbivore
86-1 716
Solitary
Temnothorax
Absent
Arb&Gro
Omnivore
50-300
Cooperative
Nesomyrmex
Absent
Arb&Gro
Omnivore
22-47
Cooperative
Orectognathus
Absent
Ground
Predator
< 104
Cooperative
Pheidole
Absent
Arb&Gro
Omnivore
22-10 000
Cooperative
Pogonomyrmex
Absent
Ground
Omnivore
21-12 358
Cooperative
Procryptocerus
Absent
Arboreal
Omnivore
62
?
Sericomyrmex
Absent
Ground
Herbivore
300-2 000
?
Stenamma
Absent
Ground
Omnivore
12-250
?
Tetramorium
Absent
Arb&Gro
Predator
10 975
Cooperative
Trachymyrmex
Absent
Ground
Herbivore
>1 000
Cooperative
Wasmannia
Absent
Arb&Gro
Omnivore
839
Cooperative
Ectatomminae (4/0/0-4)
Ectatomma
Present
Ground
Omnivore
30-400
Solitary
Gnamptogenys
Present
Arb&Gro
Predator
100-1 000
Cooperative
Rhytidoponera
Present
Ground
Omnivore
50-271
Solitary
Typhlomyrmex
Present
Ground
Predator
100-1 000
?
Heteroponerinae (1/0/0-3)
Heteroponera
Present
Ground
Omnivore
30-100
?
1Data of cocoon state mainly come from (Armitage et al., 2012) and (Greer & Moreau, 2021). 2 Data of nesting habit, diet, mean colony size and foraging behavior come from (Greer & Moreau, 2021) and (Blanchard et al., 2016). *Subfamily division and the number of genera per subfamily are referenced from Antwiki (AntWiki Website). Brackets under subfamily name indicate the number of genera, whose pupae are covered/polymorphic/naked, as well as the total genus number in this subfamily.
Diagnosis: Presence of peg-like teeth on labrum and clypeus; mandibles long, strongly curved with a long apical tooth and a strong subapical tooth; eyes present; geniculate antenna with twelve articles, with a long scape; propodeal lobes well developed; petiole well defined, pedunculate, with a rather large node and a strong constriction between it and gaster (Nel, et al., 2004).
Figure
1.
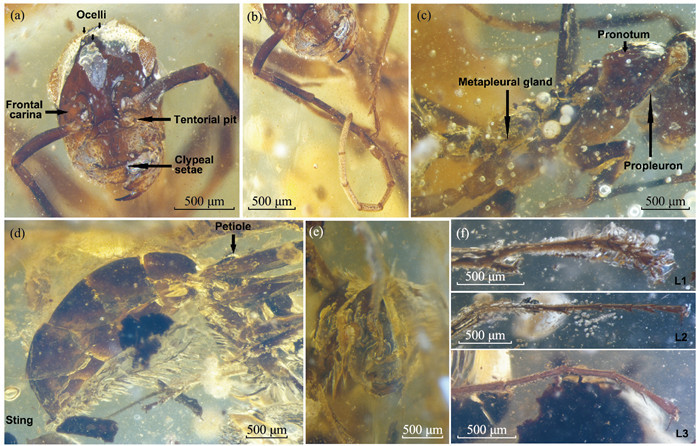
Photomicrographs of Gerontoformica sp. in the Myanmar amber specimen STJ-452 Holotype (Individual 1): (a) Frontal view of head features; (b) Frontal view of right antenna; (c) Dorsolateral view ofmesosoma. Paratype (Individual 2): (d) Lateralview of metasoma; (e) Frontal view of head features; (f) Details of left legs’ tarsus and claw.
Diagnosis: Workers, similar to other Gerontoformica species. Distinguishing features include the presence of antennal scrobes, head elongate, 1.44× longer than wide, frontal lobe absent; 20-22 denticles running along the anterior margin of the clypeus, and ocelli small (0.07 mm wide). The body length was approximately 5.06 mm.
Description: Head: Head 1.28 mm long (anterior margin of clypeus to posterior margin of head), head 0.89 mm wide (below eyes). No apparent microsculpture on cuticle of head. Mandible simple, with only two teeth; outer surface with numerous, widely scattered, fine setae. Clypeus length 0.21 mm along the medial axis, sparse tapered setae present on dorsal surface; anterior margin of the clypeus with 20-22 peglike denticles, clypeal posterior margin with three lobes. Ocelli present, small, front ocellus 0.07 mm in diameter, two setae inside the two posterior ocelli. Occiput rounded, oval shaped. Eye present, 0.32 mm long and 0.22 mm wide, inner margins 0.68 mm apart. Antennal scrobes, oval shaped. Torulus and bulbus exposed. Antenna with 12 segments, total length 4.49 mm: scape 0.67, pedicel 0.23, funicular segment I 0.56, II 0.37, III 0.30, IV 0.29, V 0.25, VI 0.35, VII 0.32, VIII 0.33, IX 0.33, X 0.49 mm. Mesosoma: Segment lengths, from lateral view: pronotum 1.02 mm, mesonotum 0.41 mm, metanotum 0.40 mm, propodeum 0.49 mm, suture hardly visible owing to shelter by bubble inclusions and fissures. Pronotum-propleuron sulci well-developed; propleuron reduced, still visible in lateral view.Propodeum gradually rounded, propodeal spiracle turret-like. Legs very long, with very sparse setae. Procoxa 0.19 mm long, 0.25 mm wide; protrochanter 0.09 mm long, 0.26 mm wide; profemur 1.46 mm long, 0.24 mm wide; protibia 1.18 mm long, 0.09 mm wide, with one pectinate and one simple tibial spur equal in length. Mesocoxa 0.60 mm long, 0.27 mm wide; mesotrochanter 0.26 mm long, 0.16 mm wide; mesofemur 1.74 mm long, 0.21 mm wide; mesotibia 1.46 mm long, 0.08 mm wide, two mesotibial spurs present, one pectinate, the other simple and slightly shorter. Metacoxa 0.53 mm long, 0.21 mm wide; metatrochanter 0.17 mm long, 0.24 mm wide; metafemur 1.96 mm long, 0.27 mm wide; mesotibia 1.49 mm long, 0.14 mm wide, two mesotibial spurs present, one trick and pectinate, one thin and simple. Tarsi with five tarsomeres, all segments setose, with four thick setae per segment at apex of each tarsomere.Pretarsal claws with subapical teeth. Metasoma: Petiole nodiform, with short ened peduncle and rounded dorsal node.Propodeal-petiolar attachment height 0.16 mm. Petiole length 0.52 mm in lateral view.Node gradually rounded, height 0.32 mm, shortened to 0.12 mm at gaster attachment point. Gastral segment I (abdominal segment III) length 0.37 mm, II 0.59 mm, III 0.48 mm, IV 0.39 mm, V 0.53 mm. Sting extruded and broken, retained part 0.11 mm long (For describing details see Fig. 1).
Morphology of Cocoons
3 cocoons were preserved in the amber specimen STJ-452. All had similar textures and morphologies, and thus were obviously spun by the same species, Gerontoformica sp. Two worker ant individuals of which were also preserved in this amber. Individual one showed a light-colored exoskeleton and a curled-up posture, which suggests that it was probably captured by the resin soon after its emergence (Fig. 2a). The exoskeleton of individual two was darker than that of individual one; thus, we identified it as an older individual who emerged for a period of time, probably an adult remover ant in the nest (Fig. 2b). In addition, the splayed legs of individual two with a large number of bubbles on their periphery also suggest that it was a highly active individual; these bubbles are usually caused by struggling when the insects are captured in resin.
Figure
2.
Cretaceous worker ants and empty cocoons in the specimen STJ-452:(a) Ventral view of worker individual one (W1) and cocoon one (C1);(b) Lateral view of worker individual two (W2);(c) Photomicrograph of cocoon two (C2) and three (C3);(d) Front end of C3, showing the emergence hole under transmission light; (e) Tail end of C3, showing the meconium under transmission light; (f) Texture of the C2.
Among these cocoons, two were preserved completely, with a length of approximately 59 mm× 20 mm. All cocoons were empty and oriented along the direction of the resin flow, surrounded by woody debris and insect frass (Fig. 2a-2c). As with many extant crowd-group ants, the cocoons of Gerontoformica sp. were translucent and membranous in texture, with a brown coloration observed through the amber. The morphology of the two integrated cocoons showed that the front end of the cocoons was rounded, while the tail end was elongated backward and had accumulated meconium (Fig. 2d and Fig. 2e) (Sarzetti, et al., 2019). Under the stereoscope, the surface of cocoons was wrinkled, and the silk fibers could be distinguished (Fig. 2f). The cocoons have small stomas on their surfaces, especially on the tail end (Fig. 2f). Near the front end of these cocoons, there was a large irregular hole, which was supposed to be caused by the emergence of the ant (Fig. 2d).
The second specimen, STJ-446, preserved a hymenopteran within its cocoon (Fig. 3a). It measured 85 mm in length and 28 mmin width. We identified it as a stem-group ant based on the cocoon texture, which is similar to that of Gerontoformica sp. (Fig. 3d), as well as its explicit petiole and sting (Fig. 3b and Fig. 3c). In addition, the amber had captured some dipsters and a tumbling flower beetle, as in (Bao, et al., 2019).
Figure
3.
Cretaceous ant pupa in cocoon, preserved in specimen STJ-446:(a)Lateral view of the pupa; (b) Pygidium in detail, showing the sting and the third valvula; (c) Petiole in detail; (d) Texture of the cocoon.
Phylogenetic Analysis and Ancestral State Reconstruction
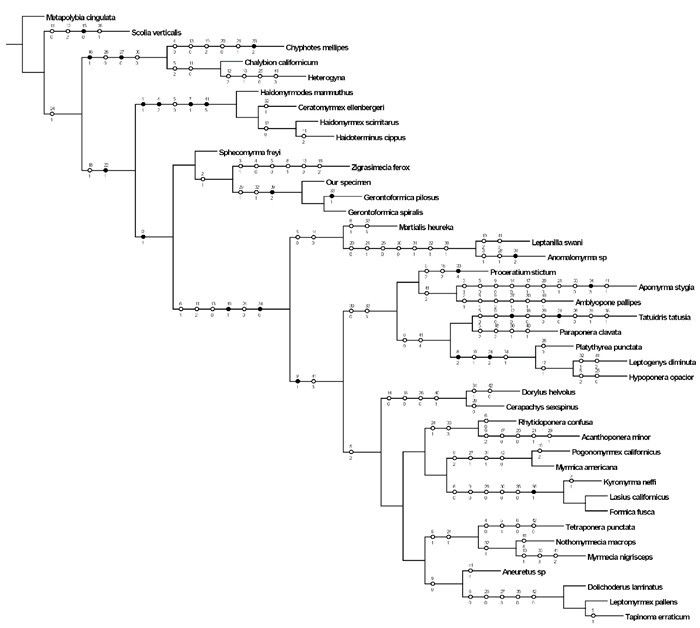
The “traditional” and “new technology” searches, under equal or implied weighting, generated four strict consensus trees. Among them, we chose the tree produced by the traditional search under implied weighting to conduct subsequent analysis, because this search is more suitable for small datasets and implied weighting can lead to more stable topologies and improved resampling support (Goloboff et al., 2010). After constraining the crown group with the topological structure of the tree produced by molecular data (Borowiec et al., 2019), a strict consensus tree and comparison of the main characters with phylogenetic relevance are shown in Fig. 4 (length = 214, consistency index = 31, retention index = 60).
Figure
4.
Strict consensus tree yielded by Traditional Search (Equal-weight) in TNT v. 1.5.Unambiguous morphological character states are shown on the tree with a black circle as the homologous state and a white circle as the homoplasious state.
Consistent with previous research, the cladistic analysis of Gerontoformica sp. and other stem-group ants (Sphecomyrminae and Haidomyrmecinae) places them as paraphyletic taxa to all extant ants. Meanwhile, Haidomyrmecinae was supported as a monophyletic subfamily, and was placed as sister to all remaining ants. Our phylogenetic analysis showed a Haidomyrmecinae + (Sphecomyrminae + other living subfamilies) pattern. However, probably because of the morphological diversity of extant ants, some crown-group ant subfamilies are unsatisfactorily clustered together.
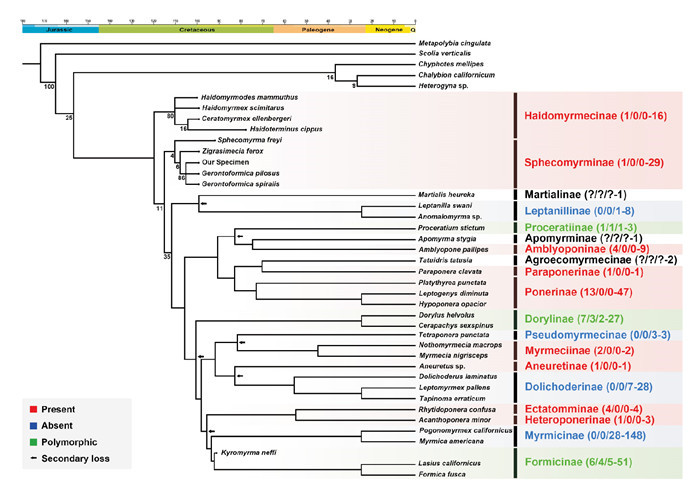
Using ancestral character state reconstruction, we found that cocoon-covered pupae are plesiomorphic in Formicidae, even though the existence of cocoons is unstable. Leptanillinae, Dolichoderinae, Pseudomyrmecinae and Myrmicinae have a naked pupal stage, whereas Proceratiinae, Dorylinae and Formicinae show a polymorphic pattern. Furthermore, our results also indicate that the cocoon-spinning behavior was lost at least six times in Formicidae: (1) the common ancestor of Leptanillinae; (2) the interior of Proceratiinae; (3) the interior of Dorylinae; (4) the common ancestor of Dolichoderinae; (5) the common ancestor of Pseudomyrmecinae; (6) the common ancestor of Myrmicinae and (partial) Formicinae (Fig. 5).
Figure
5.
Phylogenetic tree of Formicidae, marked with presence and absence of pupal cocoons on the subfamily level.
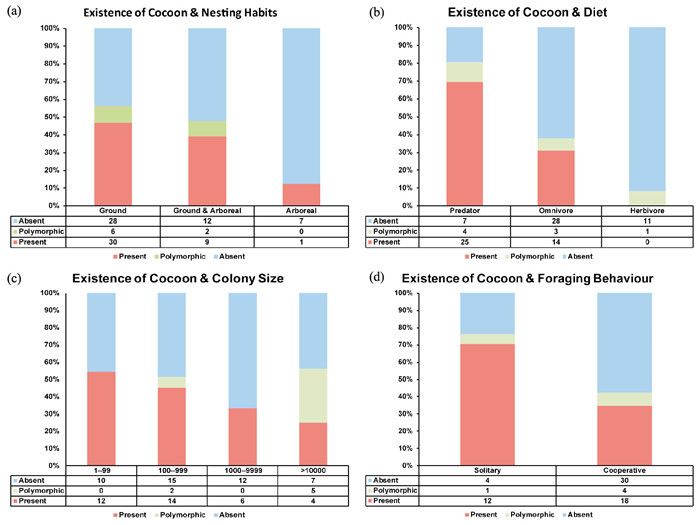
The statistical analysis of the relationship between the cocoon’s state and the four factors showed similar patterns (Fig. 6). The proportion of genera with a covered pupal stage decreases with the transition of nesting habit from the ground (46.9%) to ground and arboreal (39.1%), and finally to arboreal (12.5%). Regarding diet, the figures for predators, omnivores and herbivores make up 69.4%, 31.1%, and 0% respectively. The percentage of genera with cocoons also declined with an increase in the mean colony size, from 1-99 (54.5%) to more than 10 000 (25%). Meanwhile, regarding the switch in foraging behavior, the percentage with cocoons falls from 70.6% for solitary to 34.6% for cooperative.
Figure
6.
Proportions of the genus with documented cocoon existence, showing the relationship between the existence of cocoons and four factors: (a) nesting habits, (b) diet, (c) colony size, and (d) foraging behavior. For data see Table 2.
Ancestral Origin of Cocoon-Spinning Behavior in Ants
The life cycle of holometabolous insects usually has four stages, which means that between the larval and adult stages, there is a sessile period known as the pupal stage.During this period, because of their vulnerability to natural enemies, many endopterygota insects would develop an extensive diversity of methods, such as chemical toxins, physical structure (spines, cocoons, protective coloration), and the recruitment of other species to guard them, to aid them through this immobile stage (Lindstedt et al., 2019). A wide variety of insect pupa fossils and bio-sedimentary structures have been reported (Brasier et al., 2017). However, fossil records of these protective behaviors or structures are rare, probably owing to poor preservation conditions or research bias.
So far, cocoon-like fossils have been found in Argentina, England and Myanmar, with the time spanning from the Early Cretaceous to the Late Eocene. For example, six possible insect cocoons were found in southern England (Early Cretaceous, ca. 140 Ma), and these cocoons were coated with resin, which suggests that their producers may use plant resin to cover their cocoons to deter predators (Brasier et al., 2017). Several calcareous cocoon-like trace fossils have been reported from the lacustrine-palustrine limestone formation in southern England (Late Eocene), and they were interpreted as non-hymenopteran insect chambers for pupation, aestivation, or hibernation (Edwards et al., 1998). 3 ichnofossils of insect pupation chambers were described from Patagonia and Argentina (Cretaceous), some of which were attributable to wasp cocoons based on their similarity in morphological structures (Genise et al., 2007). 8 wasp cocoon fossils were preserved within a broken dinosaur egg in Argentina (Late Cretaceous, 70-80 Ma), which indicates that these wasp larvae fed on dried organic matter and span cocoons in it (Genise & Sarzetti, 2011). Last but not least, 3 workers and 1 pupa of stem ant Gerontoformica gracilis were reported from Myanmar amber recent year (Boudinot et al., 2022). While in this study, we report additional 2 ant cocoon specimens from Myanmar amber. Compared with compacted fossils, cocoons preserved in amber can be observed from different directions and present more microstructures of the cocoon surface and the internal insect. Therefore, they can provide a reference for the comparative study of the Mesozoic cocoon structures, which can help to identify the sources of empty cocoons preserved in amber specimens or compacted fossils in future research.
Among the Hymenoptera, which is one of the largest insect orders on Earth, the ability to produce silk mainly used for the construction of nests and cocoons has evolved several times (Sutherland et al., 2012). Thus, in recent years, to determine the evolutionary process of this behavior in the order Hymenoptera, some researchers have mapped cocoon characteristics (morphologies, compositions, and textures) onto a Hymenoptera phylogeny tree (Sarzetti et al., 2019). The results showed that spinning silk to produce cocoons is plesiomorphic in Hymenoptera, building membranous cocoons can already be seen in some basal lineages, such as Tenthredinoidea and Ichneumonidae (Sarzetti et al., 2019). However, the phylogenetic evolutionary patterns of cocoon characters in the different clades of Hymenoptera are still poorly understood.
Ants belong to the Aculeate subclade of Hymenoptera, and it is generally assumed that theyare closely related to wasps and bees (Wilson et al., 1967). Like many other Hymenoptera members, many species of Formicidae also produce silk cocoons before the pupation stage to isolate themselves from outside environments (Armitage et al., 2012). Analysis of the “fibroin” proteins of aculeates (bees, wasps, and ants) suggests that these paralogous silk proteins are derived from duplications of a single sequence, which arose before the divergence of bees from ants and wasps over 150 million years ago (Campbell et al., 2014). Thus, it can be assumed that spinning silk to construct cocoons before pupation appeared in the ancestors of ants. While our specimens represent the most basal lineage of ants — Sphecomyrminae, they had silk cocoons during the pupal stage, which supports the hypothesis that producing a cocoon is plesiomorphic of Formicidae.
Secondary Loss of Cocoons in Ants
According to previous research, cocoons can play an important role in protecting their occupants from environmental damage or attack by natural enemies; however, there are still some ant clades that lack this characteristic. In Myrmicinae, fungus-growing ants (Attini), who develop an obligate mutualism with fungi cultivated in their nest, would use the fungal mycelium to cover their pupae instead of silk cocoons (Armitage et al., 2012). In addition, some other ant species in Formicinae might even have a mixed pupa stage, with both covered and naked pupae in their nests (Purcell & Chapuisat, 2012). Therefore, we collected data on cocoons from all kinds of sources and used them to reconstruct the ancestral state of the cocoon-spinning behavior at subfamily level. The results show that this ancient behavior was lost at least six times in Formicidae (Table 2, Fig. 5).
Relative relationships among 27 crown-group ant species from 17 extant subfamilies, 9 stem-group ant species from 2 extinct subfamilies, and 5 outgroup representatives. The analysis was conducted with 43 morphological characters mainly from (Barden & Grimaldi, 2016), using a “traditional” search (equal-weighting), and eventually constrained by the extent of the topological structure and divergence dates from (Borowiec et al., 2019). Nodes and branch lengths donot correspond to the actual diversification times ormorphological changes. The node values represent the bootstrap support measures. Note added in proof: ant cocoons produced by Haidomyrmecinae found in Myanmar amber (Fig. 7).
Figure
7.
Ant cocoons produced by subfamily Haidomyrmecinae found in Myanmar amber (Pictures provided by the amber’s owner).
To explain the secondary loss of the cocoon, some researchers ascribed it to the nesting habit of ants (Armitage et al., 2012). Unlike social wasps and bees, ants usually build nests in soil or rotten woods, where environmental factors, such as temperature and humidity, are easier to control; thus, they do not need a cocoon to regulate these environmental factors (Armitage et al., 2012). However, our results showed the opposite trend. Reviewing the evolutionary history of ants, these widespread insects were rare in the Mesozoic. The large-scale molecular phylogenetic analysis suggested that ants originated from the Middle Jurassic to Early Cretaceous, but began to diversify during the Late Cretaceous to Early Eocene (Moreau et al., 2006). This period of time is consistent with the rise of angiosperm forests, in which diverse leaf litter and abundant herbivorous insects provided a wider array of habitats and direct or indirect foods to ants, respectively (Moreau et al., 2006). Therefore, during the expansion of angiosperm forests, primitive ants started to forage on trees and gradually nest on them, which has been supported by the previous ancestral state reconstruction of ant nesting and feeding habits (Blanchard et al., 2016; Nelsen et al., 2018). During this time, our statistical analysis shows that as ants’ nesting habits transform from ground to arboreal, their feeding habits change from being direct predators to indirect herbivores, the percentage of taxa at the genus level whose pupa are covered by silk cocoons decreases steadily (Fig. 6a and Fig. 6b). This result suggests that ground nesting and direct-hunting taxa are more likely to spin cocoons before the pupal stage, probably to defend against pathogens in the complex microbial environment of soil or leaf litter.
However, some researchers claim that cocoon-spinning behavior is related to the development of brood care. Previous research shows that if cocoons are naked, it is more convenient for worker ants to groom and monitor their pupae in the nest (Armitage et al., 2012). In addition, the energy costs and time consumption to form a cocoon, which delay the development process of the ant individual for 2.3 days on average, also not make it worth it to produce a cocoon in safe environments (Purcell & Chapuisat, 2012).All these factors can be attributed to eusocial evolution. In the current world, the four dominant ant subfamilies are Ponerinae, Dolichoderinae, Formicinae and Myrmicinae (Wilson & Bert, 2005). Among them, compared with the other three families, Ponerinae are socially primitive, characterized by small colony sizes, poor brood care, and the lone foraging of worker ants (Wilson & Bert, 2005). Our statistical analysis showed that all Ponerinae have a covered pupal stage. Moreover, as the colony size increases and the transition from solitary foraging to cooperative foraging is undergone, the proportion of genus with silk cocoons also shows a marked decline (Fig. 6c and Fig. 6d). This implies that with the development of eusociality, ants are inclined to have a naked pupal stage, possibly because of the need for rapid colony expansion and more efficient brood care. Overall, our statistical results show a similar pattern in four factors, and one conclusion can be put forward: with the transition of nesting habits (ground to arboreal), diet (from predator to omnivore, and finally shifting to herbivore), and the development of eusociality (characterized by larger colony sizes and cooperative foraging) in ants, their cocoons are steadily lost; however, which factor plays the dominant role in this decline is still unclear.
Taphonomy and Eusociality in Stem-Group Ants
Both amber specimens contained some wood particulates and insect frass, which indicates that they were formed on or near the forest floor (Perrichot, 2004). However, in their extant counterparts, ant larvae and pupae are usually assembled in covert nests. How were these pupae captured in the resin? For specimen STJ-446, this amber contains an ant pupa within its cocoon, a tumbling flower beetle (Bao et al., 2019), and dipsters. Therefore, we suggest that this specimen was formed in an exposed environment, probably on the ground of the forest floor. The ant pupa may have been dropped accidently by the remover ant and become trapped in the resin.
Regarding the assemblage of empty cocoons and worker ants in specimen STJ-452, we gave 3 possibilities for its taphonomy: (1) the worker ants moved the pupae underground and were captured in resin by accident, similar to specimen STJ-446. However, this hypothesis cannot explain why all of the pupae were empty; it is unreasonable for the ants to carry these empty pupae outside of the nest. (2)Gerontoformica sp. lived in a half-open nest beside or under the resin-producing trees, and the resin flowed into the ant nest and eventually formed this amber specimen. (3) The empty cocoons and newly hatched worker ant’s corpse were moved out of the nest, and discarded in the waste disposal zones of the ant colony, where they became embedded in the resin. In the third explanation, individual two may be a remover, and individual one may already have been dead before it was moved out of the nest. This interpretation can explain the difference in the mobility of the two ant individuals, and why these empty cocoons, worker ants, and dipsters were captured in the same amber specimen together. Thus, we recommend this interpretation. If that is the case, it would be the first time that corpse and garbage removal behavior is described in a society of stem-group ants (Meunier, 2015).
Thus far, it is generally assumed that Cretaceous stem-group ants were already eusocial, supported by the caste differentiation, mating flights and many morphological features adapted to social life (Barden & Grimaldi, 2016). However, according to the definition of eusociality, as well as caste differentiation, the ants should also meet 2 other important requirements: overlapping generations and brood care by worker ants in the nest (Breed et al., 1998). However, until now, these two characteristics have been seldom reported. There is only one record of brood caring in a mid-Cretaceous ant (Anderson, 2009); in that Myanmar amber specimen, a worker ant of undetermined subfamily was preserved with some suspected eggs. This worker was assigned to a basal lineage of Ponerinae based on its stout body and small eyes (Anderson, 2009). However, we argue that the overall appearance and eye size are variable in Sphecomyrminae (Phillip & David, 2014). Based on its mouthpart and antenna structures, we suggest that worker ants in the original paper should be redistributed into Sphecomyrminae. Therefore, if the identification of the ant eggs in that paper is correct, this will be—to the best of our knowledge—the first to report brood care in stem-group ants. At this time, the worker ants preserved with undisputed cocoons may further support the hypothesis that brood care has been acquired by Sphecomyrminae, providing evidence for eusociality in stem-group ants.
CONCLUSION
Numerous holometabolous insects would spin cocoons before their pupal stage, which can protect them from predators, pathogens, and fluctuant environmental factors in the external environment. It is generally assumed that the cocoon-spinning behavior is plesiomorphic in Hymenoptera. However, the phylogenetic evolutionary patterns of this behavior in the different clades of Hymenoptera (e.g. Formicidae) are still poorly understood. Herein, we describe two ant pupae specimens from Myanmar amber. Our finding supports the ancestral origin of cocoon-spinning behavior in Formicidae. In addition, based on phylogenetic analysis and ancestral state reconstruction, this behavior was secondarily lost at least six times in ants, which could be attributed to the transition of ant nesting and feeding habits, as well as the development of eusociality. Finally, empty cocoons preserved with work ants simultaneously suggest garbage removal or brood care behavior in stem-group ants, improving our understanding of their eusociality in the Cretaceous.
ACKNOWLEDGMENTS
We are grateful for the help on ant identification from Professor Zhi-lin Chen (Guangxi Normal University). We are also thankful to other researchers in the laboratory of evolution crossing domains for their helpful discussions.
Anderson, S. R. (2009). A primitive ant brood chamber with evidence of brood care in Burmese amber (Lower Cretaceous) - implications for brood care as the facilitating factor for true eusociality and dominance of ants. Denisia, (26): 11-20.
Armitage, S. A. O., Fernandez-Marin, H., Wcislo, W. T., et al. (2012). An evaluation of the possible adaptive function of fungal brood covering by Attine ants. Evolution, 66(6): 1 966-1 975. doi: 10.1111/j.1558-5646.2011.01568.x
[4]
Bao, T., Wang, B., Li, J., Dilcher, D. (2019). Pollination of Cretaceous flowers. Proceedings of the National Academy of Sciences of the United States of America, 116(49), 201916186.
[5]
Barden, P., Grimaldi, D. A. (2016). Adaptive radiation in socially advanced stem-group ants from the Cretaceous. Current Biology, 26(4): 515-521. doi: 10.1016/j.cub.2015.12.060
[6]
Barden, P. (2017). Fossil ants (Hymenoptera: Formicidae): Ancient diversity and the rise of modem lineages. Myrmecological News, 24: 1-30.
[7]
Blanchard, B. D., Moreau, C. S. (2016). Defensive traits exhibit an evolutionary trade-off and drive diversification in ants. Evolution, 71(2): 315-328.
[8]
Bolton, B. (2003). Synopsis and classification of Formicidae (vol. 71). Memoirs of the American Entomological Institute, The American Entomological Institute, Gainesville.
Borowiec, M., Rabeling, C., Brady, S., et al. (2019). Compositional heterogeneity and outgroup choice influence the internal phylogeny of the ants. Molecular Phylogenetics and Evolution, 134: 111-121. doi: 10.1016/j.ympev.2019.01.024
[11]
Boudinot, B., Richter, A., Katzke, J., et al. (2022). Evidence for the evolution of eusociality in stem ants and a systematic revision of Gerontoformica (Hymenoptera: Formicidae). Zoological Journal of the Linnean Society, 195(4): 1 355-1 389. doi: 10.1093/zoolinnean/zlab097
[12]
Brady, S. G., Schultz, T. R., Fisher, B. L., et al. (2006). Evaluating alternative hypotheses for the early evolution and diversification of ants. Proceedings of the National Academy of Sciences, 103(48): 18 172-18 177. doi: 10.1073/pnas.0605858103
[13]
Brasier, A. T., Cotton, L. J., Garwood, R. J., et al. (2017). Earliest Cretaceous cocoons or plant seed structures from the Wealden Group, Hastings, UK. In A. T. Brasier, D. McIlroy, & N. McLoughlin (Eds.), Earth System Evolution and Early Life: A Celebration of the Work of Martin Brasier, 448: 399-411.
[14]
Breed, M., Choe, J., Crespi, B. (1998). The evolution of social behavior in Insects and Arachnids. Bioscience, 48(1): 61-62. doi: 10.2307/1313230
[15]
Campbell, P. M., Trueman, H. E., Zhang, Q., et al. (2014). Cross-linking in the silks of bees, ants and hornets. Insect Biochemistry and Molecular Biology, 48: 40-50. doi: 10.1016/j.ibmb.2014.02.009
[16]
Cremer, S., Armitage, S. A. O., Schmid-Hempel, P. (2007). Social immunity. Current Biology, 17(16): R693-R702. doi: 10.1016/j.cub.2007.06.008
[17]
Cruickshank, R. D., Ko, K. (2003). Geology of an amber locality in the Hukawng Valley, Northern Myanmar. Journal of Asian Earth Sciences, 21(5): 441-455. doi: 10.1016/S1367-9120(02)00044-5
[18]
Edwards, N., Jarzembowski, E. A., Pain, T., et al. (1998). Cocoon-like trace fossils from the lacustrine-palustrine Bembridge Limestone Formation (Late Eocene), Southern England. Proceedings of the Geologists Association, 109: 25-32. doi: 10.1016/S0016-7878(98)80003-5
[19]
Genise, J. F., Melchor, R. N., Bellosi, E. S., et al. (2007). New insect pupation chambers (Pupichnia) from the Upper Cretaceous of Patagonia, Argentina. Cretaceous Research, 28(3): 545-559. doi: 10.1016/j.cretres.2006.08.007
[20]
Genise, J. F., Sarzetti, L. C. (2011). Fossil cocoons associated with a dinosaur egg from Patagonia, Argentina. Palaeontology, 54(4): 815-823. doi: 10.1111/j.1475-4983.2011.01064.x
Goloboff, P. A., Carpenter, J. M., Arias, J. S., et al. (2010). Weighting against homoplasy improves phylogenetic analysis of morphological data sets. Cladistics, 24(5), 758-773.
[23]
Greer, J., Moreau, C. (2021). Phylogenetic analysis and trait evolution of ant cocoons. Insect Systematics & Evolution, 53(1): 1-18.
[24]
Grimaldi, D. A., Engel, M. S., Nascimbene, P. C. (2002). Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. American Museum Novitates, 62(3 361), : 1-71.
[25]
Krynicki, V. E. (2013). Primitive ants (Hymenoptera: Sphecomyrminae) in the Campanian (Late Cretaceous) of North Carolina (USA). Life: The Excitement of Biology, 1(3): 156-165. doi: 10.9784/LEB1(3)Krynicki.03
[26]
Latreille, P. A. (1809). Genera crustaceorum et insectorum secundum ordinem in familias disposita (vol. 4). Parisiis et Argentorati: A. Koenig.
[27]
LaPolla, J. S., Dlussky, G. M., Perrichot, V. (2013). Ants and the fossil record. In M. R. Berenbaum (Ed.), Annual Review of Entomology, 58: 609-630. doi: 10.1146/annurev-ento-120710-100600
[28]
Lindstedt, C., Murphy, L., Mappes, J. (2019). Antipredator strategies of pupae: How to avoid predation in an immobile life stage?Philosophical Transactions of the Royal Society B-Biological Sciences, 374(1 783): 20190069.
[29]
Maddison, W., Maddison, D. (2009). Mesquite: a modular system for evolutionary analysis. Evolution, 11(5).
[30]
Meunier, J. (2015). Social immunity and the evolution of group living in insects. Philosophical Transactions of the Royal Society B-Biological Sciences, 370(1 669), 10: 20140102.
[31]
Moreau, C. S., Bell, C. D., Roger, V., et al. (2006). Phylogeny of the ants: Diversification in the age of angiosperms. Science, 312(5 770): 101-104.
[32]
Nel, A., Perrault, G., Perrichot, V., et al. (2004). The oldest ant in the lower cretaceous amber of Charente-maritime (SW France)(Insecta: Hymenoptera: Formicidae). Geologica Acta, 2(1): 23-30.
[33]
Nelsen, M. P., Ree, R. H., Moreau, C. S. (2018). Ant-plant interactions evolved through increasing interdependence. Proceedings of the National Academy of Sciences of the United States of America, 115(48): 12 253-12 258.
[34]
Perrichot, V. (2004). Early Cretaceous amber from south-western France: Insight into the Mesozoic litter fauna. Geologica Acta, 2(1), 9-22.
[35]
Perrichot, V., Wang, B., Barden, P. J. C. R. (2020). New remarkable hell ants (Formicidae: Haidomyrmecinae stat. nov.) from mid-Cretaceous amber of northern Myanmar. Cretaceous Research, 109: 104 381.
[36]
Peters, R. S., Krogmann, L., Mayer, C., et al. (2017). Evolutionary History of the Hymenoptera. Current Biology, 27(7): 1 013-1 018. doi: 10.1016/j.cub.2017.01.027
[37]
Phillip, B., David, G. (2014). A diverse ant fauna from the mid-cretaceous of Myanmar (Hymenoptera: Formicidae). Plos One, 9(4): e93 627. doi: 10.1371/journal.pone.0093627
[38]
Pull, C., Ugelvig, L., Wiesenhofer, F., et al. (2018). Destructive disinfection of infected brood prevents systemic disease spread in ant colonies. Elife, 7: e32 073. doi: 10.7554/eLife.32073
[39]
Pull, C. D., Metzler, S., Naderlinger, E., et al. (2018). Protection against the lethal side effects of social immunity in ants. Current Biology, 28(19): R1 139-R1 140. doi: 10.1016/j.cub.2018.08.063
[40]
Purcell, J., Chapuisat, M. (2012). The influence of social structure on brood survival and development in a socially polymorphic ant: insights from a cross-fostering experiment. Journal of Evolutionary Biology, 25(11): 2 288-2 297. doi: 10.1111/j.1420-9101.2012.02607.x
[41]
Sarzetti, L. C., Genise, J. F., Dinghi, P., et al. (2019). An overview of hymenopteran cocoons as a tool to interpret ichnospecies of Fictovichnus (Pallichnidae) and other fossil cocoons of wasps. Palaios, 34(11): 562-574. doi: 10.2110/palo.2019.053
[42]
Shi, G., Grimaldi, D. A., Harlow, G. E., et al. (2012). Age constraint on Burmese amber based on U-Pb dating of zircons. Cretaceous Research, 37: 155-163. doi: 10.1016/j.cretres.2012.03.014
[43]
Sutherland, T. D., Weisman, S., Walker, A. A., et al. (2012). Invited review: The coiled coil silk of bees, ants, and hornets. Biopolymers, 97(6): 446-454. doi: 10.1002/bip.21702
[44]
Tragust, S., Ugelvig, L. V., Chapuisat, M., et al. (2013). Pupal cocoons affect sanitary brood care and limit fungal infections in ant colonies. Bmc Evolutionary Biology, 13: 225. doi: 10.1186/1471-2148-13-225
[45]
Westerweel, J., Roperch, P., Licht, A., et al. (2019). Burma Terrane part of the Trans-Tethyan Arc during collision with India according to palaeomagnetic data. Nature Geoscience, 12(5-6): 863-868.
[46]
Wilson, E. O., Bert, H. L. (2005). The rise of the ants: A phylogenetic and ecological explanation. Proceedings of the National Academy of Sciences of the United States of America, 102(21): 7 411-7 414.
[47]
Wilson, E. O., Carpenter, F. M., Brown, W. L. (1967). The first Mesozoic ants, with the description of a new subfamily. Psyche: A Journal of Entomology, 74(1): 1-19. doi: 10.1155/1967/89604
Table
2.
Documentary of cocoon state on the genus level, coded with the nesting habit, diet, colony size and foraging behavior in Formicidae*
Genus
Cocoons State1
Nesting Habit2
Diet2
Mean Colony Size2
Forging Behavior2
Leptanillinae (0/0/1-8)
Leptanilla
Absent
Ground
Predator
150
Cooperative
Proceratiinae (1/1/1-3)
Discothyrea
Present
Ground
Predator
73
?
Proceratium
Polymorphic
Arb&Gro
Predator
13-200
Solitary
Probolomyrmex
Absent
Ground
Predator
20
Solitary
Amblyoponinae (4/0/0-9)
Apomyrma
Present
Ground
Predator
< 100
?
Amblyopone
Present
Ground
Predator
12
Cooperative
Mystrium
Present
Ground
Predator
< 200
Cooperative
Onychomyrmex
Present
Ground
Predator
50-400
Cooperative
Ponerinae (13/0/0-47)
Diacamma
Present
Ground
Predator
30-60
Solitary
Dinoponera
Present
Ground
Omnivore
30-60
Solitary
Harpegnathos
Present
Ground
Predator
35-225
Solitary
Anochetus
Present
Arb&Gro
Predator
100-400
Solitary
Cryptopone
Present
Ground
Predator
30-999
?
Hypoponera
Present
Ground
Predator
21-1 500
?
Leptogenys
Present
Ground
Predator
10-50 000
Cooperative
Myopias
Present
Ground
Predator
< 100
Cooperative
Odontomachus
Present
Arb&Gro
Predator
18-10 000
Cooperative
Ophthalmopone
Present
Ground
Predator
20-800
Solitary
Pachycondyla
Present
Ground
Omnivore
9-800
Cooperative
Platythyrea
Present
Arb&Gro
Predator
19-500
Solitary
Ponera
Present
Ground
Predator
30-60
Cooperative
Genus
Cocoons State1
Nesting Habit2
Diet2
Mean Colony Size2
Forging Behavior2
Paraponerinae (1/0/0-1)
Paraponera
Present
Arb&Gro
Omnivore
>2 000
Cooperative
Dorylinae (7/3/2-27)
Acanthostichus
Present
Ground
Predator
?
Solitary
Aenictus
Polymorphic
Ground
Predator
850 000
Cooperative
Cerapachys
Polymorphic
Ground
Predator
75-200
?
Dorylus
Absent
Ground
Predator
100 000
Cooperative
Eciton
Present
Ground
Predator
100 000
Cooperative
Sphinctomyrmex
Present
Ground
?
?
?
Simopone
Present
Arboreal
Predator
?
?
Syscia
Absent
Ground
?
?
?
Labidus
Present
Ground
Predator
1 000 000
Cooperative
Neivamyrmex
Polymorphic
Ground
Predator
110 000
Cooperative
Nomamyrmex
Present
Ground
Predator
1 000
Cooperative
Lioponera
Present
Arb&Gro
Predator
?
Cooperative
Aneuretinae (1/0/0-1)
Aneuretus
Present
Ground
Omnivore
65-75
Cooperative
Dolichoderinae (0/0/7-28)
Azteca
Absent
Arboreal
Omnivore
100-1 000
Cooperative
Bothriomyrmex
Absent
Ground
Omnivore
?
?
Dolichoderus
Absent
Arb&Gro
Omnivore
25-1 000
Cooperative
Iridomyrmex
Absent
Ground
Omnivore
79 000
Cooperative
Leptomyrmex
Absent
Ground
Omnivore
350
?
Linepithema
Absent
Arb&Gro
Omnivore
>1 000
Cooperative
Tapinoma
Absent
Arb&Gro
Omnivore
100-5 000
Cooperative
Myrmeciinae (2/0/0-2)
Myrmecia
Present
Ground
Omnivore
15-1 500
Cooperative
Nothomyrmecia
Present
Ground
Omnivore
50-70
Solitary
Pseudomyrmecinae(0/0/3-3)
Myrcidris
Absent
Arboreal
Herbivore
100
?
Pseudomyrmex
Absent
Arb&Gro
Omnivore
200-10 000
Solitary
Tetraponera
Absent
Arboreal
Omnivore
7 000
Cooperative
Formicinae (6/4/5-51)
Acropyga
Polymorphic
Ground
Herbivore
?
?
Brachymyrmex
Present
Arb&Gro
Omnivore
22-125
Cooperative
Camponotus
Present
Arb&Gro
Omnivore
55 000
Cooperative
Colobopsis
Absent
Arboreal
Herbivore
?
?
Formica
Polymorphic
Ground
Omnivore
150 000 000
Cooperative
Gesomyrmex
Absent
Arboreal
Omnivore
150
?
Gigantiops
Present
Ground
Omnivore
>1 000
Solitary
Lasius
Polymorphic
Ground
Omnivore
1 250 000
Cooperative
Myrmoteras
Present
Ground
Predator
8-22
Solitary
Myrmecocystus
Present
Ground
Omnivore
15 000
?
Oecophylla
Absent
Arboreal
Omnivore
500 000
Cooperative
Paratrechina
Absent
Arb&Gro
Omnivore
2 000
Cooperative
Plagiolepis
Present
Arb&Gro
Omnivore
< 5 000
?
Polyrhachis
Polymorphic
Arb&Gro
Omnivore
60-90 000
?
Prenolepis
Absent
Ground
Omnivore
3 370
Cooperative
Myrmicinae (0/0/28-148)
Acromyrmex
Absent
Arb&Gro
Herbivore
1 000-10 000
Cooperative
Aphaenogaster
Absent
Ground
Omnivore
100-300
Cooperative
Apterostigma
Absent
Ground
Herbivore
30-150
Cooperative
Atta
Absent
Ground
Herbivore
1 750 000
Cooperative
Cardiocondyla
Absent
Ground
Omnivore
>50
Cooperative
Crematogaster
Absent
Arb&Gro
Omnivore
40-29 000
Cooperative
Cyphomyrmex
Absent
Ground
Herbivore
< 500
Solitary
Eutetramorium
Absent
Ground
Predator
1-50
Cooperative
Formicoxenus
Absent
Ground
Omnivore
< 100
?
Harpagoxenus
Absent
Ground
Omnivore
10
Cooperative
Leptothorax
Absent
Ground
Omnivore
20-60
Cooperative
Genus
Cocoons State1
Nesting Habit2
Diet2
Mean Colony Size2
Forging Behavior2
Messor
Absent
Ground
Herbivore
100-1 000
Cooperative
Monomorium
Absent
Ground
Omnivore
>10 000
Cooperative
Mycocepurus
Absent
Ground
Herbivore
< 1 000
?
Myrmecina
Absent
Ground
Omnivore
24-100
?
Myrmica
Absent
Ground
Predator
86-3 000
?
Myrmicocrypta
Absent
Ground
Herbivore
86-1 716
Solitary
Temnothorax
Absent
Arb&Gro
Omnivore
50-300
Cooperative
Nesomyrmex
Absent
Arb&Gro
Omnivore
22-47
Cooperative
Orectognathus
Absent
Ground
Predator
< 104
Cooperative
Pheidole
Absent
Arb&Gro
Omnivore
22-10 000
Cooperative
Pogonomyrmex
Absent
Ground
Omnivore
21-12 358
Cooperative
Procryptocerus
Absent
Arboreal
Omnivore
62
?
Sericomyrmex
Absent
Ground
Herbivore
300-2 000
?
Stenamma
Absent
Ground
Omnivore
12-250
?
Tetramorium
Absent
Arb&Gro
Predator
10 975
Cooperative
Trachymyrmex
Absent
Ground
Herbivore
>1 000
Cooperative
Wasmannia
Absent
Arb&Gro
Omnivore
839
Cooperative
Ectatomminae (4/0/0-4)
Ectatomma
Present
Ground
Omnivore
30-400
Solitary
Gnamptogenys
Present
Arb&Gro
Predator
100-1 000
Cooperative
Rhytidoponera
Present
Ground
Omnivore
50-271
Solitary
Typhlomyrmex
Present
Ground
Predator
100-1 000
?
Heteroponerinae (1/0/0-3)
Heteroponera
Present
Ground
Omnivore
30-100
?
1Data of cocoon state mainly come from (Armitage et al., 2012) and (Greer & Moreau, 2021). 2 Data of nesting habit, diet, mean colony size and foraging behavior come from (Greer & Moreau, 2021) and (Blanchard et al., 2016). *Subfamily division and the number of genera per subfamily are referenced from Antwiki (AntWiki Website). Brackets under subfamily name indicate the number of genera, whose pupae are covered/polymorphic/naked, as well as the total genus number in this subfamily.

 DownLoad:
DownLoad: